Репликация вируса в клетке. Сборка вирусов. Высвобождение дочерних вирионов из клетки.
Обновлено: 05.05.2024
В цикле репродукции вирусов различают четыре стадии: 1) подготовительную, или инициальную, включающую фазы адсорбции вируса на клетке, проникновения и раздевания в клетке; 2) собственно репродуктивную стадию образования структурных белков и вирионных нуклеиновых кислот; 3) сборку вирионов; 4) заключительную, сопровождающуюся выходом зрелых вирусных частиц из клетки (рисунок 30).
Адсорбция.Первая стадия репродуктивного цикла — адсорбция вириона на поверхности инфицируемой клетки. Адсорбция происходит путём взаимодействия вириона со специфическими клеточными рецепторами. Понятие «тропизм вирусов» объясняется специфическим взаимодействием вирусных белков с поверхностными рецепторами инфицируемой клетки.
Процесс адсорбции протекает в две фазы: фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплементарности взаимодействующих молекул.

Рисунок 30 – Основные этапы репродукции вирусов
Количество инфекционных вирусных частиц, адсорбированных на клетке, определяет термин «множественность заражения» (инфицирования), то есть на клетке может сорбироваться большое количество вирионов. Однако, инфицированная вирусом клетка обычно толерантна к повторному заражению гомологичным вирусом.
Проникновение и «раздевание».«Голые» вирусы проникают в клетку путём эндоцитоза (виропексис[вирус + греч. pexis, прикрепление]) – погружения участка клеточной мембраны в месте их адсорбции. «Одетые» вирусы проникают в клетку путём слияния суперкапсида с клеточной мембраной при участии специфических F-белков(белков слияния). При проникновении «голых» вирусов в клетку образуются вакуоли (эндосомы). После проникновения «одетых» вирусов в цитоплазму происходит частичная депротеинизация вирионов и модификация их нуклеопротеида (раздевание). Модифицированные частицы теряют инфекционные свойства.
Теневая фаза.После депротеинизации вирусы невозможно выделить из культуры клеток. Этот этап репродукции известен как теневая фаза, или фаза эклипса[от англ. eclipse, затмение]. Она включает репликацию нуклеиновых кислот вируса и синтез вирусных белков. Теневая фаза не происходит при температуре 0−4 °С (исключая вирус гриппа). Теневая фаза заканчивается после образования составных компонентов вируса, необходимых для сборки дочерних популяций.
Сборка.У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато. Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов. В процессе образования «одетых» вирусов полные нуклеокапсиды упорядоченно выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками).
Высвобождение дочерних вирионов. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
Вопросы для самоконтроля
1 Какие формы взаимодействия вирусов с клеткой различают?
2 Какие формы продуктивности инфекции существуют?
3 Назовите стадии реплинации вирусов?
4 Охарактеризуйте начальную стадию репликации.
5 Каким способом вирусы могут проникать в клетку хозяина и от чего это зависит?
6 Как происходит синтез структурных элементов вириона?
7 Почему одну из фаз репликации вирусов называют «теневой фазой»?
8 Как происходит высвобождение дочерних популяций простых и сложных вирусов?
9 Как реализуется кодирующая стратегия вирусов в зависимости от организации генома?
Заключительные стадии репродукции вирусов. Сборка и выход вирусов из клетки
Репликацией называется синтез молекул нуклеиновой кислоты, гомологичных геному. В клетке происходит репликация ДНК, в результате которой образуются дочерние двунитчатые ДНК. Репликация происходит на расплетенных участках ДНК и идет одновременно на обеих нитях от 5'-конца к З'-концу. Поскольку две нити ДНК имеют противоположную полярность, а участок репликации («вилка») движется в одном направлении, одна цепь строится в обратном направлении отдельными фрагментами, которые называются фрагментами Оказаки (по имени ученого, впервые предложившего такую модель). После синтеза фрагменты Оказаки «сшиваются» лигазой в единую нить.
Репликация ДНК осуществляется ДНК-полимеразами. Для начала репликации необходим предварительный синтез короткого участка РНК на матрице ДНК, который называется затравкой. С затравки начинается синтез нити ДНК, после чего РНК быстро удаляется с растущего участка.
Репликация вирусных ДНК. Репликация генома ДНК-содержащих вирусов в основном катализируется клеточными фрагментами и механизм ее сходен с механизмом репликации клеточной ДНК.
Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. Такой механизм репликации называется полуконсервативным.
У вирусов, содержащих кольцевые двунитчатые ДНК (паповавирусы), разрезается одна из нитей ДНК, что ведёт к раскручиванию и снятию супервитков на определенном участке молекулы.
При репликации однонитчатых ДНК (семейство парвовирусов) происходит образование двунитчатых форм, которые представляют собой промежуточные репликативные формы.
Репликация вирусных РНК. В клетке нет ферментов, способных осуществить репликацию РНК. Поэтому ферменты, участвующие в репликации, всегда вирусспецифические. Репликацию осуществляет тот же фермент, что и транскрипцию; репликаза является либо модифицированной транскриптазой, либо при репликации соответствующим образом модифицируется матрица.
Репликация однонитчатых РНК осуществляется в два этапа: вначале синтезируются комплементарные геному нити, которые в свою очередь становятся матрицами для синтеза копий генома. У «минус-нитевых» вирусов первый этап репликации — образование комплементарных нитей сходен с процессом транскрипции. Однако между ними есть существенное отличие: если при транскрипции считываются определенные участки генома, то при репликации считывается весь геном. Например, иРНК парамиксовирусов и рабдовирусов являются короткими молекулами, комплементарными разным участкам генома, а иРНК вируса гриппа на 20—30 нуклеотидов короче каждого фрагмента генома. В то же время матрицы для репликации являются полной комплементарной последовательностью генома и называются антигеномом. В зараженных клетках существует механизм переключения транскрипции на репликацию. У «минус-нитевых» вирусов этот механизм обусловлен маскировкой точек терминации транскрипции на матрице генома, в результате чего происходит сквозное считывание генома. Точки терминации маскируются одним из вирусных белков.
При репликации растущая «плюс-нить» вытесняет ранее синтезированную «плюс-нить» либо двухспиральная матрица консервируется. Более распространен первый механизм репликации.
Репликативные комплексы. Поскольку образующиеся нити ДНК и РНК некоторое время остаются связанными с матрицей, в зараженной клетке формируются репликативные комплексы, в которых осуществляется весь процесс репликации (а в ряде случаев также и транскрипции) генома. Репликативный комплекс содержит геном, репликазу и связанные с матрицей вновь синтезированные цепи нуклеиновых кислот. Вновь синтезированные геномные молекулы немедленно ассоциируются с вирусными белками, поэтому в репликативных комплексах обнаруживаются антигены. В процессе репликации возникает частично двунитчатая структура с однонитчатыми «хвостами», так называемый репликативный предшественник (РП).
Репликативные комплексы ассоциированы с клеточными структурами либо с предсуществующими, либо вирусиндуцируемыми. Например, репликативные комплексы пикорнавирусов ассоциированы с мембранами эндоплазматической сети, вирусов оспы — с цитоплазматическим матриксом, репликативные комплексы аденовирусов и вирусов герпеса в ядрах находятся в ассоциации со вновь сформированными волокнистыми структурами и связаны с ядерными мембранами. В зараженных клетках может происходить усиленная пролиферация клеточных структур, с которыми связаны репликативные комплексы, или их формирование из предсуществующего материала. Например, в клетках, зараженных пикорнавирусами, происходит пролиферация гладких мембран. В клетках, зараженных реовирусами, наблюдается скопление микротрубочек; в клетках, зараженных вирусами оспы, происходит формирование цитоплазматического матрикса.
В репликативных комплексах одновременно с синтезом геномных молекул осуществляется транскрипция и происходит сборка нуклеокапсидов и сердцевин, а при некоторых инфекциях — и вирусных частиц. О сложной структуре репликативных комплексов говорит, например, такой состав репликативного комплекса аденовирусов: реплицирующиеся ДНК, однонитчатые ДНК, однонитчатые РНК, ферменты репликации и транскрипции, структурные и неструктурные вирусные белки и ряд клеточных белков.
Регуляция репликации. Вновь образованная молекула геномной РНК может быть использована различным образом. Она может ассоциироваться с капсидными белками и войти в состав вириона, служить матрицей для синтеза новых геномных молекул, либо — для образования иРНК, наконец, у «плюс-нитевых» вирусов она может выполнять функции иРНК и связываться с рибосомами. В клетке существуют механизмы, регулирующие использование геномных молекул. Регуляция идет по принципу саморегуляции и реализуется путем взаимодействия вирусных РНК и белков благодаря возможности белокнуклеинового и белок-белкового узнавания. Например, роль терминального белка пикорнавирусов заключается в запрещении трансляции иРНК и отборе молекул для формирования вирионов. Белок, связывающийся с 5'-концом геномной РНК, в свою очередь узнается капсидными белками и служит сигналом для сборки вирусной частицы с участием данной молекулы РНК. По тому же принципу отбираются геномные молекулы РНК у «минус-нитевых» вирусов: к З'-концу геномных РНК присоединяется молекула капсидного вирусного белка, к которой подстраиваются другие белковые субъединицы в результате белок-белкового узнавания, и такая молекула РНК войдет в состав вириона или послужит матрицей для репликации. Для переключения ее на транскрипцию должен возникнуть запрет белок-нуклеинового взаимодействия. В репликации ДНК аденовирусов участвует молекула белка, которая связывается с концом вирусной ДНК и необходима для начала репликации. Таким образом, для начала репликации необходим синтез вирусных белков: в присутствии ингибиторов белкового синтеза отсутствует переключение транскрипции на репликацию.
СБОРКА ВИРУСНЫХ ЧАСТИЦ
Синтез компонентов вирусных частиц в клетке разобщен и может протекать в разных структурах ядра и цитоплазмы. Вирусы, репликация которых проходит в ядрах, условно называют ядерными. В основном это ДНК-содержащие вирусы: аденовирусы, паповавирусы, парвовирусы, вирусы герпеса. Вирусы, реплицирующиеся в цитоплазме, называют цитоплазматическими. К ним относятся из ДНК-содержащих вирус оспы и большинство РНК-содержащих вирусов, за исключением ортомиксовирусов и ретровирусов. Однако это разделение весьма относительно, потому что в репродукции тех и других вирусов есть стадии, протекающие соответственно в цитоплазме и ядре.
Внутри ядра и цитоплазмы синтез вирусспецифических молекул также может быть разобщен. Так, например, синтез одних белков осуществляется на свободных полисомах, а других — на полисомах, связанных с мембранами. Вирусные нуклеиновые кислоты синтезируются в ассоциации с клеточными структурами вдали от полисом, которые синтезируют вирусные белки. При таком дисъюнктивном способе репродукции образование вирусной частицы возможно лишь в том случае, если вирусные нуклеиновые кислоты и белки обладают способностью при достаточной концентрации узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и самопроизвольно соединяться друг с другом, т. е. способны к самосборке.
В основе самосборки лежит специфическое белок-нуклеиновое и белок-белковое узнавание, которое может происходить в результате гидрофобных, солевых и водородных связей, а также стерического соответствия. Белок-нуклеиновое узнавание ограничено небольшим участком молекулы нуклеиновой кислоты и определяется уникальными последовательностями нуклеотидов в некодирующей части вирусного генома. С этого узнавания участка генома вирусными капсидными белками начинается процесс сборки вирусной частицы. Присоединение остальных белковых молекул осуществляется за счет специфичеческих белокбелковых взаимодействий или неспецифических белокнуклеиновых взаимодействий.
В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки.
1. У просто устроенных вирусов формируются провирионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато. Сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек.
2. Сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазматических вирусов — с участием мембран эндоплазматической сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы.
3. У ряда сложно устроенных вирусов существуют специальные гидрофобные белки, выполняющие функции посредников между сформированными нуклеокапсидами и вирусными оболочками. Такими белками являются матриксные белки у ряда «минус-нитевых» вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов).
4. Сборка нуклеокапсидов, сердцевин, провирионов и вирионов происходит не во внутриклеточной жидкости, а в специальных структурах, предсуществующих в клетке или индуцированных вирусом («фабриках»).
5. Сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 — гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе ареновирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот.
Сборка РНК-содержащих вирусов. Сборка просто устроенных РНК-содержащих вирусов заключается в ассоциации вирусного генома с вирусными капсидными белками с образованием нуклеокапсида.
У сложно устроенных РНК-содержащих вирусов процессы сборки нуклеокапсидов, сердцевин и зрелых вирионов обычно разобщены. Нуклеокапсиды мигрируют к месту сборки вирусных частиц — плазматической мембране (или мембранам эндоплазматическои сети) и упорядочение выстраиваются под участками мембран, с наружной стороны которых уже встроены вирусные суперкапсидные белки. Сборка заключается в том, что участки, содержащие гликопротеиды с примыкающими к ним нуклеокапсидами, постепенно выпячиваются через модифицированную клеточную мембрану. В результате выпячивания образуется «почка», содержащая нуклеокапсид и оболочку с суперкапсидными белками. «Почка» отделяется от клеточной мембраны с образованием свободной вирусной частицы. Такой способ формирования вирусных частиц называется почкованием. Почкование может происходить через плазматическую мембрану клетки в наружную среду, как у ортомиксовирусов, парамиксовирусов, рабдовирусов и альфа-вирусов, либо через мембраны эндоплазматическои сети в вакуоли, как у аренавирусов и буньявирусов, В основе выпячивания почки через мембрану лежат обычные клеточные процессы, направленные на отторжение непригодного для клетки материала и обновление мембран. Участок будущей почки содержит фиксированный нуклеокапсид, ассоциированный с суперкапсидными белками, но движение мембранных липидов продолжается в силу их текучести, липиды обволакивают будущую почку и вместе с ними из «почки» вытесняются клеточные мембранные белки. В результате этого движения происходит выбухание «почки» над клеточной мембраной. Механизм образования «почки» объясняет, почему в составе почкующихся вирусов не содержится клеточных мембранных белков.
Все вирусные компоненты — нуклеокапсиды и суперкапсидные белки прибывают к месту сборки независимо друг от друга. Первыми к месту сборки прибывают суперкапсидные белки. Обычно этими белками являются гликопротеиды, которые синтезируются в полисомах, связанных с мембранами, и через шероховатые, а затем гладкие мембраны в результате слияния с ними везикул комплекса Гольджи транспортируются на наружную поверхность плазматических мембран или остаются в составе везикул.
Включение гликопротеидов в определенные зоны клеточных мембран приводит к модификациям мембран. Нуклеокапсид узнает эти участки и подходит к ним с внутренней стороны липидного бислоя. Узнавание осуществляется с помощью одного из двух механизмов, 1) нуклеокапсид взаимодействует с участком гликопротеида, пронизывающим клеточную мембрану и вышедшим на ее внутреннюю поверхность. Такой механизм имеет место у альфа-вирусов; гидрофобный фрагмент гликопротеида Е1 проникает через липидный слой на его внутреннюю поверхность, и с этим фрагментом связываются нуклеокапсиды, которые позже войдут в состав «почки»; 2) в сборку вовлекается еще один вирусный белок, являющийся медиатором сборки, который называется мембранным, или матриксным белком. М-белок синтезируется на свободных полисомах, но сразу после синтеза встраивается в клеточные мембраны с внутренней цитоплазматической стороны липидного бислоя. Этот белок в высокой степени гидрофобен и поэтому способен к белок-белковым и белоклипидным взаимодействиям.
Включение М-белка в клеточные мембраны является сигналом для сборки вирусной частицы: вслед за включением немедленно следует связывание нуклеокапсидов с мембранами и почкование вирусной частицы. Тем самым М-белок обладает функцией лимитирующего сборку фактора.
Сборка ДНК-содержащих вирусов. В сборке ДНК-содержащих вирусов есть некоторые отличия от сборки РНК-содержащих вирусов. Как и у РНК-содержащих вирусов, сборка ДНК-содержащих вирусов является многоступенчатым процессом с образованием промежуточных форм, отличающихся от зрелых вирионов по составу полипептидов. Первый этап сборки заключается в ассоциации ДНК с внутренними белками и формировании сердцевин или нуклеокапсидов. При этом ДНК соединяется с предварительно сформированными «пустыми» капсидами.
В результате связывания ДНК с капсидами появляется новый класс промежуточных форм, которые называются неполными формами. Помимо неполных форм с разным содержанием ДНК, существует другая промежуточная форма в морфогенезе — незрелые вирионы, отличающиеся от зрелых тем, что содержат ненарезанные предшественники полипептидов. Таким образом, морфогенез вирусов тесно связан с модификацией (процессингом) белков.
Сборка ядерных вирусов начинается в ядре, обычно — с ассоциации с ядерной мембраной. Формирующиеся в ядре промежуточные формы вируса герпеса почкуются в перинуклеарное пространство через внутреннюю ядерную мембрану, и вирус приобретает таким путем оболочку, которая является дериватом ядерной мембраны. Дальнейшая достройка и созревание вирионов происходит в мембранах эндоплазматической сети и в аппарате Гольджи, откуда вирус в составе цитоплазматических везикул транспортируется на клеточную поверхность.
У непочкующихся липидсодержащих вирусов — вирусов оспы сборка вирионов происходит в уже описанных цитоплазматических вирусных «фабриках». Липидная оболочка вирусов в «фабриках» формируется из клеточных липидов путем автономной самосборки, поэтому липидный состав оболочек значительно отличается от состава липидов в клеточных мембранах.
ВЫХОД ВИРУСНЫХ ЧАСТИЦ ИЗ КЛЕТКИ
Существуют два способа выхода вирусного потомства из клетки: 1) путем «взрыва»; 2) путем почкования.
Выход из клетки путем взрыва связан с деструкцией клетки, нарушением ее целостности, в результате чего находящиеся внутри клетки зрелые вирусные частицы оказываются в окружающей среде. Такой способ выхода из клетки присущ вирусам, не содержащим липопротеидной оболочки (пикорна-, рео-, парво-, папова-, аденовирусы). Однако некоторые из этих вирусов могут транспортироваться на клеточную поверхность до гибели клетки.
Выход из клеток путем почкования присущ вирусам, содержащим липопротеидную мембрану, которая является дериватом клеточных мембран. При этом способе клетка может длительное время сохранять жизнеспособность и продуцировать вирусное потомство, пока не произойдет полное истощение ее ресурсов.
Репликация вируса в клетке. Сборка вирусов. Высвобождение дочерних вирионов из клетки.
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.

Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
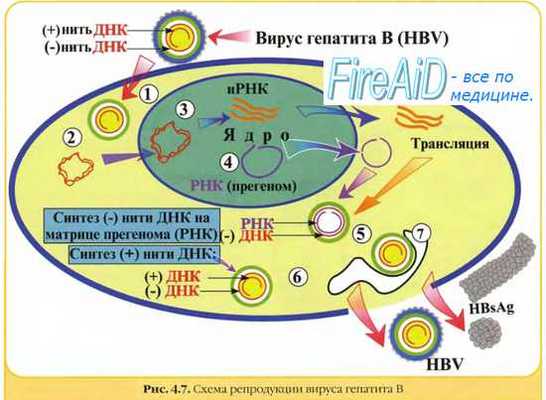
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Латентное инфицирование клеток вирусам. ДНК некоторых вирусов (герпесвирусы, ретровирусы) может находиться в клетке вне хромосом, либо вирусная ДНК интегрируется в ядерный геном, но вирусспецифические синтезы не происходят. Такая вирусная ДНК образует латентный провирус, реплицирующийся вместе с хромосомой. Подобные состояния вирусной ДНК нестабильны, возможны периодические реактивации с переходом в продуктивное взаимодействие «вирус-клетка», либо клетка трансформируется, давая начало злокачественному росту.

Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.

Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
Первая стадия репродуктивного цикла — адсорбция вириона на поверхности инфицируемой клетки. Адсорбция происходит путём взаимодействия вириона со специфическими клеточными рецепторами. За распознавание рецепторов ответственны белки, входящие в состав капсида либо суперкапсида. Таким образом, понятие «тропизм вирусов» объясняется специфическим взаимодействием вирусных белков с поверхностными рецепторами инфицируемой клетки. Например, полиовирус проникает в клетки центральной нервной системы (ЦНС) и желудочно-кишечного тракта (ЖКТ) и размножается в них, так как у человека и приматов только эти клетки имеют рецепторы к белкам полиовирусов,
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
• Количество инфекционных вирусных частиц, адсорбированных па клетке, определяет термин «множественность заражений» (инфицирования). Обычно животная клетка содержит около 50 000 рецепторов, и её заражение носит множественный характер, то есть на клетке может сорбироваться большое количество вирионов. Тем не менее инфицированная вирусом клетка обычно толерантна к повторному заражению гомологичным вирусом.
В зависимости от типа генетического материала (ДНК или РНК), образование дочерних копий геномов протекает по-разному.
У ДНК-геномных вирусов репликация вирусных ДНК принципиально сходна с репликацией клеточных ДНК.
Репликацию РНК-геномных вирусов осуществляют вирусные РНК-зависимые РНК-полимеразы (репликазы). Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
Сборка вирусов
У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато.
Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов, или сердцевин. В процессе образования «одетых - вирусов полные нуклеокапсиды упорядочение выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками). При нарушениях процесса самосборки могут образовываться пустые капсиды либо комплексы нуклеиновых кислот с внутренними белками.

Высвобождение дочерних вирионов из клетки
Высвобождение дочерних вирионов — конечная стадия репродуктивного цикла. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны (ЦПМ) и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. Принцип высвобождения дочерних популяций почкованием во многом сходен с процессами, направленными на отторжение непригодного для клетки материала или обновление клеточных мембран. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
- Вернуться в оглавление раздела "Микробиология."
Читайте также: