Возбуждение клетки. Возникновение потенциала действия в клетке
Обновлено: 19.05.2024
Потенциалом действия (ПД) называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения. Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды.
В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе. Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы.
В ПД выделяют следующие фазы (рис. 3):

Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала. Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 - +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации.
Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов - следовую деполяризацию и следовую гиперполяризацию.
Ионный механизм возникновения ПД. Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К + из цитоплазмы во внешний раствор превышает противоположно направленный поток Na + . Поэтому наружная сторона мембраны в покое имеет положительный потенциал по отношению к внутренней.
При действии на клетку раздражителя проницаемость мембраны для Na + резко повышается и становится примерно в 20 раз больше проницаемости для K + . Поэтому поток Na + из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутренняя сторона мембраны в месте возбуждения становится заряженной положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе ПД (фазе деполяризации).
Повышение проницаемости мембраны для Na + продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na + вновь понижается, а для K + возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией. В результате инактивации поток Na + внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K + из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД.
ПД возникает при деполяризации поверхностной мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению ионов Na внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные изменения на мембране (локальный ответ). При увеличении силы раздражения, когда достигнут порог возбудимости, изменения мембранного потенциала достигают критического уровня деполяризации (КУД). Например, величина потенциала покоя равна -70 мВ, КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т.е. на -20 мВ снизить ее исходный потенциал покоя. Только при достижении КУД наблюдается резкое изменение мембранного потенциала, которое регистрируется в виде ПД. Таким образом, основное условие возникновения потенциала действия - это снижение мембранного потенциала до критического уровня деполяризации.
В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами:
избирательностью (селективностью) по отношению к определенным ионам;
электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала.
Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны.
Активный и пассивный ионный транспорт. В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na + и K + по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии.
Таким образом, в живой клетке существует две системы движения ионов через мембрану.
Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны.
Второй осуществляется против концентрационного градиента. Он состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт - результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения.
Потенциал покоя и потенциал действия
Мембрана всех живых клеток поляризована. Внутренняя сторона мембраны несет отрицательный заряд по сравнению с межклеточным пространством (рис. 1). Величина заряда, который несет мембрана называется мембранным потенциалом (МП). В невозбудимых тканях МП низкий, и составляет около -40 мВ. В возбудимых тканях он высокий, около -60 - -100 мВ и называется потенциалом покоя (ПП).
Потенциал покоя, как и любой мембранный потенциал формируется за счет избирательной проницаемости клеточной мембраны. Как известно, плазмолемма состоит из липидного бислоя, через который движение заряженных молекул затруднено. Белки, встроенные в мембрану, могут избирательно изменять проницаемость мембраны для различных ионов, в зависимости от приходящих стимулов. При этом, для формирования потенциала покоя ведущую роль играют ионы калия, кроме них важны ионы натрия и хлора.
Рис. 1. Концентрации и распределение ионов с внутренней и внешней стороны мембраны.
Большинство ионов распределяются неравномерно с внутренней и внешней стороны клетки (рис. 1). Внутри клетки концентрация ионов калия выше, а натрия и хлора - ниже, чем снаружи. В состоянии покоя мембрана проницаема для ионов калия и практически непроницаема для ионов натрия и хлора. Несмотря на то, что калий может свободно выходить из клетки, его концентрации остаются неизменными благодаря отрицательному заряду на внутренней стороне мембраны. Таким образом, на калий действуют две силы, находящиеся в равновесии: осмотические (градиент концентрации К + ) и электрические (заряд мембраны), благодаря чему число входящих в клетку ионов калия равно выходящим. Движение калия осуществляется через калиевые каналы утечки, открытые в состоянии покоя. Величину заряда мембраны, при которой ионы калия находятся в равновесии можно вычислить по уравнению Нернста:
где Ек — равновесный потенциал для К + ; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; n — валентность К + (+1), [К + н] — [К + вн] — наружная и внутренняя концентрации К + .
Если подставить в уравнение значения из таблицы на рис. 43, то мы получим величину равновесного потенциала, равную примерно -95 мВ. Это значение вписывается в диапазон мембранного потенциала возбудимых клеток. Отличия ПП разных клеток (даже возбудимых) могут возникать по трем причинам:
- отличия внутриклеточной и внеклеточной концентраций ионов калия в разных тканях (в таблице приведены данные по среднестатистическому нейрону);
- натрий-калиевая АТФаза может вносить свой вклад в значение заряда, так как она выводит из клетки 3 Na + в обмен на 2 К + ;
- несмотря на минимальную проницаемость мембраны для натрия и хлора, эти ионы все-таки могут попадать в клетки, хоть и от 10 до 100 раз хуже, по сравнению с калием.
Чтобы учесть проникновение других ионов в клетку существует уравнение Нернста-Гольдмана:
, где Еm — мембранный потенциал; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; РK , PNa и РCl — константы проницаемости мембраны для К + Na + и Сl, соответственно; [К + н], [K + вн], [Na + н], [Na + вн], [Сl — н] и [Сl — вн ]- концентрации K + , Na + и Сl снаружи (н) и внутри (вн) клетки.
Такое уравнение позволяет установить более точную величину ПП. Обычно, мембрана оказывается на несколько мВ менее поляризована, по сравнению с равновесным потенциалом для К + .
Потенциал действия (ПД) может возникать в возбудимых клетках. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то ПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет кратковременная перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной, после чего восстановится ПП. Это кратковременное изменение ПП, происходящее при возбуждении клетки называется потенциалом действия.
Возникновение ПД возможно благодаря тому, что в отличие от ионов калия, ионы натрия далеки от равновесия. Если подставить в уравнение Нернста натрий вместо калия, то мы получим равновесный потенциал, равный примерно +60 мВ. Во время ПД, происходит кратковременное увеличение проницаемости для Na + . При этом, натрий начнет проникать в клетку под действием двух сил: по градиенту концентрации и по заряду мембраны, стремясь подстроить заряд мембраны под свой равновесный потенциал. Движение натрия осуществляется по потенциал-зависимым натриевым каналам, которые открываются в ответ на смещение мембранного потенциала, после чего сами инактивируются.
Рис. 2. Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
На записи ПД выглядит как кратковременный пик (рис. 44), имеющий несколько фаз.
- Деполяризация (фаза нарастания) (рис. 44) - увеличение проницаемости для натрия из-за открытия натриевых каналов. Натрий стремится к своему равновесному потенциалу, но не достигает его, так как канал успевает инактивироваться.
- Реполяризация - возвращение заряда к величине потенциала покоя. Помимо калиевых каналов утечки здесь подключаются потенциал-зависимые калиевые каналы (активируются от деполяризации). В это время калий выходит из клетки, возвращаясь к своему равновесному потенциалу.
- Гиперполяризация (не всегда) - возникает в случаях, если равновесный потенциал по калию превышает по модулю ПП. Возвращение к ПП происходит после возвращения к равновесному потенциалу по К + .
Во время ПД происходит изменение полярности заряда мембраны. Фаза ПД, при которой заряд мембраны положителен, называется овершутом (рис. 2).
Благодаря Н-воротам инактивация канала происходит раньше, чем потенциал на мембране достигнет равновесной величины по натрию. После прекращения поступления натрия в клетку, происходит реполяризация за счет выходящих из клетки ионов калия. При этом к каналам утечки в этом случае подключаются еще и потениал-активируемые калиевые каналы. Во время реполяризации, в быстром натриевом канале быстро закрываются М-ворота. Н-ворота открываются гораздо медленнее и остаются закрытыми еще некоторое время после возвращения заряда к потенциалу покоя. Этот период принято называть периодом рефрактерности.
Рис. 3. Работа потенциал-управляемого натриевого канала.
Концентрации ионов внутри клетки восстанавливает натрий-калиевая АТФаза, которая с затратой энергии в виде АТФ откачивает из клетки 3 иона натрия и закачивает 2 иона калия.
По немиелинизированному волокну или по мембране мышцы потенциал действия распространяется непрерывно. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до порогового значения, в результате чего на соседнем участке возникает деполяризация. Главную роль в возникновении потенциала на новом участке мембраны предыдущий участок. При этом на каждом участки сразу после ПД наступает период рефрактерности, за счет которое ПД распространяется однонаправленно. При прочих равных условиях распространение потенциала действия по немиелинизированному аксону происходит тем быстрее, чем больше диаметр волокна. У млекопитающих скорость составляет 1-4 м/с. Поскольку у беспозвоночных животных отсутствует миелин, в гигантских аксонах кальмара скорость ПД может достигать 100 м/c.
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах немиелинизированных волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до порогового значения, что приводит к возникновению в них новых потенциалов действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-й, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Сальтаторное проведение увеличивает скорость проведения ПД 15-20 раз до 120 м/с.
Работа нейронов
Нервная система состоит из нейронов и глиальных клеток. Однако, главную роль в проведении и передаче нервных импульсов играют нейроны. Они получают информацию от множества клеток по дендритам, анализируют ее и передают или не передают на следующий нейрон.
Передача нервного импульса с одной клетки на другую осуществляется с помощью синапсов. Различают два основных типа синапсов: электрические и химические (рис. 4). Задача любого синапса - передать информацию с пресинаптической мембраны (мембрана аксона) на постсинаптическую (мембрана дендрита, другого аксона, мышцы или другого органа-мишени). Большинство синапсов нервной системы образуется между окончанием аксонов и дендритами, которые в области синапса образуют дендритные шипики.
Преимущество электрического синапса состоит в том, что сигнал с одной клетки на другую переходит без задержки. Кроме того, такие синапсы не утомляются. Для этого пре- и постсинаптические мембраны соединены поперечными мостиками, через которые ионы из одной клетки могут перемещаться в другую. Однако, существенным минусом такой системы является отсутствие однонаправленной передачи ПД. То есть, он может передаваться как с пресинаптической мембраны на постсинаптическую, так и наоборот. Поэтому, такая конструкция встречается достаточно редко и в основном - в нервной системе беспозвоночных.
Рис. 4. Схема строения химического и электрического синапсов.
Химический синапс весьма распространен в природе. О устроен сложнее, так как необходима система преобразования электрического импульса в химический сигнал, затем, вновь в электрический импульс. Все это приводит к возникновению синаптической задержки, которая может составить 0,2-0,4 мс. Кроме того, может произойти истощение запасов химического вещества, что приведет к утомлению синапса. Однако, такой синапс обеспечивает однонаправленность передачи ПД, что является его главным преимуществом.
Рис. 5. Схема работы (а) и электронная микрофотография (б) химического синапса.
В состоянии покоя окончание аксона, или пресинаптическое окончание, содержит мембранные пузырьки (везикулы) с нейромедиатором. Поверхность везикул заряжена отрицательно, чтобы предотвратить связывание с мембраной, и покрыта специальными белками, и принимающими участие в высвобождении везикул. В каждом пузырьке находится одинаковое количество химического вещества, которое называется квантом нейромедиатора. Нейромедиаторы весьма разнообразны по химическому строению, однако, большинство из них производятся прямо в окончании. Поэтому, в нем могут находиться системы, для синтеза химического посредника, а также аппарат Гольджи и митохондрии.
Постсинаптическая мембрана содержит рецепторы к нейромедиатору. Рецепторы могут быть в виде как ионных каналов, открывающихся при контакте со своим лигандом (ионотропные), так и мембранными белками, запускающими внутриклеточный каскад реакций (метаботропные). Один нейромедиатор может иметь несколько как ионотропных, так и метаботропных рецепторов. При этом, часть из них может быть возбуждающими, а часть - тормозными. Таким образом, реакцию клетки на нейромедиатор будет определять тип рецептора на ее мембране, и разные клетки могут совершенно по-разному реагировать на одно и то же химическое вещество.
Между пре- и постсинаптической мембраной располагается синаптическая щель, шириной 10-15 нм.
При приходе ПД на пресинаптическое окончание, на нем открываются потенциал-активируемые кальциевые каналы и ионы кальция входят в клетку. Кальций связывается с белками на поверхности везикул, что приводит к их транспортировке к пресинаптической мембране с последующим слиянием мембран. После такого взаимодействия нейромедиатор оказывается в синаптической щели (рис. 5) и может связаться со своим рецептором.
Ионотропные рецепторы - это лиганд-активируемые ионные каналы. Это значит, что канал открывается только в присутствии определенного химического вещества. Для разных нейромедиаторов это могут быть натриевые, кальциевые или хлорные каналы. Ток натрия и кальция вызывает деполяризацию мембраны, поэтому такие рецепторы называют возбуждающими. Хлорный ток приводит к гиперполяризации, что затрудняет генерацию ПД. Следовательно, такие рецепторы называют тормозными.
Метаботропные рецепторы к нейромедиаторам относят к классу рецепторов, ассоцированных с G-белками (GPCR). Эти белки запускают разнообразные внутриклеточные каскады реакций, приводящих в конечном итоге либо к дальнейшей передачи возбуждения, либо к торможению.
После передачи сигнала необходимо быстро удалить нейромедиатор из синаптической щели. Для этого в щели присутствуют либо ферменты расщепляющие, нейромедиатор, либо на пресинаптическом окончании или соседних глиальных клетках могут располагаться транспортеры, закачивающие медиатор в клетки. В последнем случае он может использоваться повторно.
Каждый нейрон получает импульсы от 100 до 100 000 синапсов. Одиночная деполяризация на одном дендрите не приведет к дальнейшей передаче сигнала. На нейрон могут приходит одновременно множество как возбуждающих, так и тормозных стимулов. Все они суммируются на соме нейрона. Такая суммация называется пространственной. Далее, может возникнуть или не возникнуть (в зависимости от пришедших сигналов) ПД в области аксонного холмика. Аксонный холмик - это область аксона, примыкающая к соме и обладающая минимальным порогом ПД. Далее импульс распространяется по аксону, конец которого может сильно ветвиться и образовывать синапсы со множеством клеток. Помимо пространственной, существует временная суммация. Она происходит в случае, поступления часто повторяющихся импульсов от одного дендрита.
Помимо классических синапсов между аксонами и дендритами или их шипиками, существуют также синапсы, модулирующие передачу в других синапсах (рис. 6). К ним относят аксо-аксональные синапсы. Такие синапсы способны усиливать или тормозить синаптическую передачу. То есть, если на окончание аксона, образующего аксо-шипиковый синапс, пришел ПД, а в это время по аксо-аксональному синапсу на него пришел тормозный сигнал, высвобождения нейромедиатора в аксо-шипиковом синапсе не произойдет. Аксо-дендритные синапсы могут изменять проведение мембраной ПД на пути от шипика к соме клетки. Также существуют аксо-соматические синапсы, которые могут влиять на суммацию сигнала в области сомы нейрона.
Таким образом, существует огромное многообразие различных синапсов, отличающихся по составу нейромедиаторов, рецепторов и их местоположению. Все это обеспечивает разнообразие реакций и пластичность нервной системы.
ЛЕКЦИЯ 2. ФИЗИОЛОГИЯ ВОЗБУЖДЕНИЯ
Раздражители и раздражимость. На живой организм постоянно действуют различные раздражители (свет, звук, различные запахи и др.). Воздействие раздражителя на организм называется раздражением. Организм воспринимает раздражение благодаря особой способности - раздражимости. Раздражимость - это способность клеток, тканей усиливать или уменьшать активность в ответ на воздействие раздражителей. Условно раздражители можно подразделить на три группы: физические, химические и физико-химические. К физическим раздражителям относятся механические, электрические, температурные, световые звуковые. К химическим относятся гормоны, лекарственные вещества и др. К физико-химическим раздражителям относятся изменения осмотического давления и рН крови.
К действию одних раздражителей орган специально приспособлен. Такие раздражители называют адекватными. Неадекватными будут такие раздражители, к воздействию которых данная клетка или ткань не приспособлена. Так для глаза адекватным раздражителем будут световые лучи, а неадекватным звуковые волны.
По силе раздражители подразделяются на подпороговые, пороговые и надпороговые. Пороговый раздражитель характеризуется минимальной силой, достаточной для того чтобы вызвать минимальный специфический эффект в раздражаемой ткани. Подпороговый раздражитель вызывает лишь местную реакцию. Его силы недостаточно для вызывания специфического эффекта. Напротив, надпороговые раздражители обладают большой силой и вызывают самую большую реакцию.
Возбудимость и возбуждение. Некоторые ткани организма (нервная, мышечная) относятся к возбудимым т. е. они обладают способностью отвечать на раздражение - возбуждением. Возбуждение - это специфическая форма реагирования возбудимой клетки или ткани на действие раздражителя. Возбуждение характеризуется как специфическими, так и неспецифическими признаками. К специфическим признакам относят сокращение мышцы, выделение железой секрета. Неспецифические признаки возбуждения - это повышение обмена веществ, усиление теплопродукции изменение электрического состояния клеточной мембраны.
Биоэлектрические явления в возбудимых тканях. Биоэлектрические явления (животное электричество) было открыто в 1791 году итальянским ученым Л. Гальвани. Современные данные происхождения биоэлектрических явлений были получены в 1952 году А. Ходжикиным, А. Хаксли и Б. Катцем в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм).
Мембранный потенциал покоя (МПП). В состоянии физиологического покоя наружная поверхность клеточной мембраны заряжена электроположительно, а внутренняя - электроотрицательно. Благодаря этому меду ними возникает разность потенциалов 60-90 мВ. Эту разность потенциалов называют мембранным потенциалом покоя (МПП) или потенциалом покоя. Возникновение потенциала покоя обусловлено неодинаковой концентрацией несущих электрические заряды ионов К , СI . В клетке преобладают крупные органические анионы, которые не могут диффендировать через мембрану.
В покое проницаемость мембраны значительно выше для К . В силу своей высокой концентрации ионы K , а внутри из отрицательно заряженных крупных анионов.
Потенциал действия (ПД). Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Na сначала медленно, а затем лавинообразно устремляется внутрь клетки. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак (деполяризация). Он становится отрицательным снаружи и положительным внутри клетки. Однако повышение проницаемости мембраны для Na . Это вызывает усиление потока положительных ионов калия из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя - отрицательный.
Волна возбуждения. Волной возбужденияназывают всю совокупность последовательных изменений электрического состояния мембраны. К компонентам волны возбуждения относятся пороговый потенциал, потенциал действия и следовые потенциалы.
Законы раздражения. В 1907 году Л. Лапик для характеристики скорости возникновения возбуждения предложил регистрировать два параметра - силу раздражения и время его воздействия. Между силой раздражения и длительностью его действия существует обратно пропорциональная зависимость: чем больше сила раздражения, тем меньше длительность его действия, необходимая для возникновения возбуждения, и наоборот. О возбудимости ткани судят по величине реобазы. Реобаза - это наименьшая сила тока (или напряжения), способная при неограниченном времени вызвать возбуждение ткани. Она измеряется в единицах силы или напряжения тока. Чем меньше реобаза, тем более возбудима ткань.
О скорости возникновения возбуждения судят по величине хронаксии. Хронаксия - это наименьшее время, в течении которого необходимо воздействовать на ткань электрическим током, равным удвоенной реобазе, чтобы вызвать ее возбуждение. Она измеряется в единицах времени. Чем меньше хронаксия тем быстрее возникает возбуждение.
Лабильность. Парабиоз. Лабильность (или функциональная подвижность ткани) - это способность возбудимой ткани к воспроизведению потенциалов действия в соответствии с ритмом раздражения. Она была открыта Н.Е. Введенским в 1892 г. Мерой лабильности является наибольшее число потенциалов действия, которое ткань способна воспроизвести в 1 с в соответствии с частотой действующих раздражителей. Лабильность является величиной непостоянной. Она может понижаться или повышаться. Понижение лабильности может наступить вследствие утомления. Повысить ее можно путем физических упражнений.
При действии на участок нерва различных факторов (солевые растворы, электрический ток, механические раздражения и т.д.) Н.Е. Введенский установил, что лабильность измененного участка понижается. Именно это состояние стойкого нераспространяющегося возбуждения Н.Е. Введенский назвал парабиозом. Парабиоз имеет три стадии. Первая стадия - уравнительная, когда и сильные и слабые раздражения, нанесенные нормальному участку нерва, вызывают одинаковое сокращение мышцы. Вторая стадия - пародоксальная, когда сильные раздражения вызывают слабое сокращения, а слабые раздражения - более сильные сокращения, чем обычно. Третья стадия - тормозящая, когда ни сильные, ни слабые раздражения не вызывают сокращения. Установленные открытия Н.Е. Введенского сыграли большую роль в дальнейшем развитии физиологии.
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Потенциалом действия называют быстрое изменение мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых железистых клеток. В основе его возникновения лежат изменения ионной проницаемости мембраны. В развитии потенциала действия выделяют четыре последовательных периода: 1) локальный ответ; 2) деполяризация; 3) реполяризация и 4) следовые потенциалы (рис. 2.11).
Локальный ответ представляет собой активную местную деполяризацию, возникающую вследствие увеличения натриевой проницаемости клеточной мембраны. Уменьшение мембранного потенциала называется деполяризацией. Однако при подпороговом стимуле начальное повышение натриевой проницаемости недостаточно велико, чтобы вызвать быструю деполяризацию мембраны. Локальный ответ возникает не только при подпороговом, но и при надпороговом раз-

Рис. 2.11. Потенциал действия:
1 — локальный ответ; 2 — фаза деполяризации; 3 — фаза реполяризации; 4 — отрицательный следовой потенциал; 5 — положительный (гиперполяризационный) следовой потенциал
дражении и является составным компонентом потенциала действия. Таким образом, локальный ответ является первоначальной и универсальной формой реагирования ткани на различные по силе раздражения. Биологический смысл локального ответа состоит в том, что если раздражитель по силе мал, то ткань реагирует на него с минимальной тратой энергии, не включая механизмы специфической деятельности. В том же случае, когда раздражение надпороговое, локальный ответ переходит в потенциал действия. Период от начала раздражения до начала фазы деполяризации, когда локальный ответ, нарастая, снижает мембранный потенциал до критического уровня (КУД), называется латентным или скрытым периодом, продолжительность которого зависит от силы раздражения (рис. 2.12).
Фаза деполяризации характеризуется быстрым уменьшением мембранного потенциала и даже перезарядкой мембраны: внутренняя ее часть на некоторое время становится заряженной положительно, а внешняя — отрицательно. Изменение знака заряда на мембране называют извращением — реверсией потенциала. В отличие от локального ответа скорость и величина деполяризации не зависят от силы раздражителя. Продолжительность фазы деполяризации в нервном волокне лягушки составляет около 0,2—0,5 мс.
Продолжительность фазы реполяризации составляет 0,5—0,8 мс. Восстановление исходной величины поляризации мембраны называют реполяризацией. В течение этого времени мембранный потен-

Рис. 2.12. Потенциалы действия, возникающие в ответ на пороговое раздражение коротким (А) и длительным (Б) стимулами Раздражающие стимулы, при воздействии которых получены ответы А и Б: ПП — потенциал покоя; Екуд. — критический уровень деполяризации мембраны (по А.Л. Каталымову)
циал постепенно восстанавливается и достигает 75—85% потенциала покоя. В литературе второй и третий периоды часто называют пиком потенциала действия.
Колебания мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов — следовую деполяризацию и следовую гиперполяризацию, которые соответствуют четвертой и пятой фазам потенциала действия. Следовая деполяризация (отрицательный следовой потенциал) является продолжением фазы реполяризации и характеризуется более медленным (по сравнению с фазой реполяризации) восстановлением потенциала покоя. Следовая деполяризация переходит в следовую гиперполяризацию (положительный следовой потенциал), представляющую собой временное увеличение мембранного потенциала выше исходного уровня. Увеличение мембранного потенциала называется гиперполяризацией. В миелинизирован- ных нервных волокнах следовые потенциалы имеют более сложный характер: следовая деполяризация может переходить в следовую гиперполяризацию, затем иногда возникает новая деполяризация, лишь после этого происходит полное восстановление потенциала покоя.
Ионный механизм возникновения потенциала действия. Основу потенциала действия составляют последовательно развивающиеся во времени изменения ионной проницаемости клеточной мембраны.
При действии на клетку раздражителя проницаемость мембраны для ионов Na + резко повышается за счет активации (открывания) натриевых каналов.
При этом ионы Na + по концентрационному градиенту интенсивно перемещаются из вне во внутриклеточное пространство. Вхождению ионов Na + в клетку способствует и электростатическое взаимодействие. В итоге проницаемость мембраны для Na + становится в 20 раз больше проницаемости для ионов К + .
Сначала деполяризация идет сравнительно медленно. Когда мембранный потенциал уменьшается на 10—40 мВ, скорость деполяризации резко увеличивается и кривая потенциала действия круто поднимается верх. Уровень мембранного потенциала, при котором резко увеличивается скорость деполяризации мембраны, благодаря тому что поток ионов Na + внутрь клетки оказывается большим, чем поток ионов К + наружу, называют критическим уровнем деполяризации.
Поскольку поток Na + в клетку начинает превышать калиевый ток из клетки, происходит постепенное снижение потенциала покоя, приводящее к реверсии — изменению знака мембранного потенциала. При этом внутренняя поверхность мембраны становится электроположительной по отношению к ее внешней электроотрицательной поверхности. Указанные изменения мембранного потенциала соответствуют восходящей фазе потенциала действия (фазе деполяризации).
Мембрана характеризуется повышенной проницаемостью для ионов Na + лишь очень короткое время (0,2—0,5 мс). После этого проницаемость мембраны для ионов Na + вновь понижается, а для К + — возрастает. В результате поток Na + внутрь клетки резко ослабляется, а ток К + из клетки усиливается.
В течение потенциала действия в клетку поступает значительное количество Na + , а ионы К + покидают клетку. Восстановление клеточного ионного баланса осуществляется благодаря работе натрий- калиевого насоса, активность которого возрастает при повышении внутренней концентрации ионов Na + и увеличении внешней концентрации ионов К + . Благодаря работе ионного насоса и изменению проницаемости мембраны для Na + и К + их концентрация во внутри- и внеклеточном пространстве постепенно восстанавливается.
Итогом этих процессов и является реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к внешней поверхности мембраны.
Следовой отрицательный потенциал регистрируется в период, когда № + -каналы инактивированы и реполяризация, связанная с выходом ионов К + из клетки, происходит медленнее, чем во время нисходящей части пика потенциала действия. Это длительное сохранение отрицательности наружной поверхности возбужденного участка по отношению к невозбужденному называют следовой деполяризацией. Следовая деполяризация означает, что в этот период наружная поверхность возбудимого образования имеет меньший положительный заряд, чем в состоянии покоя.
Следовой положительный потенциал соответствует периоду увеличения мембранного потенциала покоя, т.е. гиперполяризации мембраны. Во время следового положительного потенциала наружная поверхность клетки более положительно заряжена, чем в состоянии покоя. Следовой положительный потенциал часто называют следовой гиперполяризацией. Она объясняется длительным сохранением повышенной проницаемости для ионов К + . Вследствие этого на мембране устанавливается потенциал, равный потенциалу равновесия (для К + — 90 мВ).
Изменения возбудимости в процессе развития возбуждения. Воздействуя раздражителями разной силы в различные фазы потенциала действия, можно проследить, как изменяется возбудимость в ходе возбуждения. На рис. 2.13' видно, что период локального ответа характеризуется повышенной возбудимостью (мембранный потенциал приближается к критическому уровню деполяризации); во время фазы деполяризации мембрана утрачивает возбудимость (клетка становится рефрактерной), которая постепенно восстанавливается в ходе реполяризации.
Выделяют период абсолютной рефрактерности, который в нервных клетках продолжается около 1 мс и характеризуется их полной невоз- будимостью. Период абсолютной рефрактерности возникает в результате практически полной инактивации (непроницаемости) натриевых каналов и повышения калиевой проводимости мембраны. Даже в состоянии покоя активированы не все каналы мембраны, 40% из них находятся в состоянии инактивации. При деполяризации количество инактивированных каналов увеличивается и вершина пика потенциала действия соответствует инактивации всех натриевых каналов.
По мере реполяризации мембраны происходит реактивация натриевых каналов. Это период относительной рефрактерности: потенциал действия может возникнуть только при действии более сильных (надпороговых) раздражителей.
В период отрицательного следового потенциала фаза относительной рефрактерности сменяется фазой повышенной (супернормальной) возбудимости. В этот период порог раздражения снижен по сравнению с исходным значением, поскольку мембранный потенциал ближе к критической величине, чем в состоянии покоя (рис. 2.14) [1] [2] .
Фаза следовой гиперполяризации, обусловленная остаточным выходом калия из клетки, напротив, характеризуется снижением

Рис. 2.13. Изменение возбудимости в процессе потенциала действия:
А — компоненты волны возбуждения: 1 —деполяризация; 2 — реполяризация; МП — мембранный потенциал; мВ — микровольт; МК — критический уровень деполяризации: а — длительность порогового потенциала; б — длительность потенциала действия; в — следовая отрицательность; г — следовая положительность; Б — изменения возбудимости в разные фазы волны возбуждения; УВ — уровень возбудимости в покое: а — повышение возбудимости в период порогового потенциала; б — падение возбудимости до нуля во время протекания потенциала действия (абсолютная рефрактерность); в, — возвращение возбудимости к исходному уровню во время следовой отрицательности (относительная рефрактерность); в2 — повышение возбудимости в период конца следовой отрицательности (экзальтация'или супернормальность); в — весь период следовой отрицательности; г — падение возбудимости в период гиперполяризации (субнормальность)
возбудимости. Поскольку мембранный потенциал больше, чем в состоянии покоя, требуется более сильный раздражитель для его «смещения» до уровня критической деполяризации.
Таким образом, в динамике возбудительного процесса изменяется способность клетки реагировать на раздражители, т.е. возбудимость.

Рис. 2.14. Величина мембранного и порогового потенциалов в разные фазы развития возбуждения
Величина мембранного потенциала: Е0 — в покое; — в фазе экзальтации; Е2 — в фазе гиперполяризации. Величина порогового потенциала: е0 — в покое; е, — в фазе экзальтации; е2 — в фазе гиперполяризации
Это имеет большое значение, поскольку в момент наибольшего возбуждения (пика потенциала действия) клетка становится абсолютно невозбудимой, что защищает ее от гибели и повреждений.
Возбуждение как активная реакция клетки на раздражитель
Реакция клетки на внешнее воздействие (раздражение) отличается от реакции небиологических систем следующими особенностями:
• энергией для реакции клетки служит не энергия раздражителя, а энергия, образующаяся в результате метаболизма в самой биологической системе;
• сила и форма реакции клетки не определяется силой и формой внешнего воздействия (если сила раздражителя выше пороговой).
В некоторых специализированных клетках реакция на раздражитель проявляется особенно интенсивно. Такую интенсивную реакцию называют возбуждением.
2. Возбуждение
Возбуждение - реакция живой клетки на раздражение, выработанная в процессе эволюции: активная реакция специализированных (возбудимых) клеток на внешнее воздействие, проявляющаяся в том, что клетка начинает выполнять присущие ей специфические функции.
При возбуждении живая система переходит из состояния относительного физиологического покоя к деятельности (например, сокращение мышечного волокна, выделение секрета железистыми клетками и др.).
В организме существует 3 типа возбудимых клеток:
• нервные клетки (возбуждение проявляется генерацией электрического импульса);
• мышечные клетки (возбуждение проявляется сокращением);
• секреторные клетки (возбуждение проявляется выбросом в межклеточное пространство биологически активных веществ).
Возбудимая клетка может находиться в двух дискретных состояниях:
• состоянии покоя (готовность к реагированию на внешнее воздействие, совершение внутренней работы);
• состоянии возбуждения (активное выполнение специфических функций, совершение внешней работы).
В настоящее время достаточно хорошо изучены механизмы формирования мембранного потенциала покоя и мембранного потенциала действия, т.е процесса возбуждения клетки.
Сущность процесса возбуждения: все клетки организма имеют электрический заряд, обеспечиваемый неодинаковой концентрацией анионов и катионов вне и внутри клетки. При действии раздражителя на клетку возбудимой ткани изменяется проницаемость ее мембраны, вследствие чего ионы быстро перемещаются согласно электрохимическому градиенту (совокупность концентрационного и электрического градиентов), - это и есть процесс возбуждения. Его основой является потенциал покоя.
1) Мембранный потенциал покоя а. Общая характеристика, причина формирования
Потенциал покоя (ПП) - разность элетрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина в нервных клетках составляет 60-80 мВ. При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом скачком отклоняется и показывает отрицательный заряд внутри клетки (рисунок 1).

ПП играет исключительно важную роль в жизнедеятельности самого нейрона и организма в целом. В частности, он составляет основу для переработки информации в нервной клетке, обеспечивает регуляцию деятельности внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. Согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц), непосредственной причиной формирования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки (таблица 1).
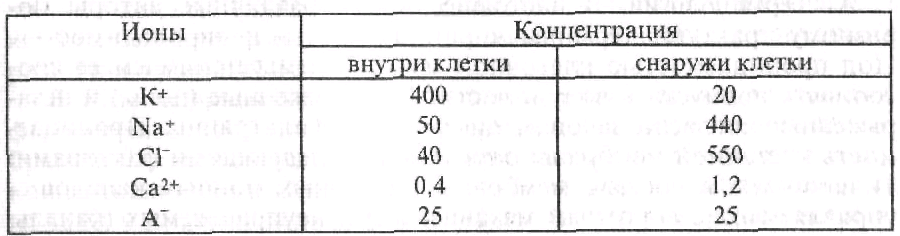
Таблица 1. Концентрация ионов в аксоплазме гигантского аксона кальмара и в экстрацеллюлярной среде в миллимолях на 1 л.
В нейронах концентрация ионов К + внутри клетки в 20-40 раз больше, чем вне клетки; концентрация ионов N3+ вне клетки в 10-12 раз больше, чем в клетке. Ионов СГ вне клетки в 10-20 раз больше, чем внутри клетки. В клетке имеется небольшое количество ионов Са 2+ . Ион Са 2+ в клетке находится в основном в связанном состоянии с АТФ, цитратом, глутаматом. Резервуаром для ионов Са 2 + является эндоплазматический ретикулум.
В свободном состоянии кальций находится в основном вне клетки; в гиалоплазме его очень мало. Это обусловлено отчасти активным транспортом ионов Са 2+ наружу через клеточную мембрану, отчасти поглощением его эндоплазматическим ретикулумом и другими органеллами, например митохондриями.
В клетке находятся также крупномолекулярные анионы, главным образом это отрицательно заряженные белковые молекулы, например глутамат, аспартат, а также органические фосфаты. Различные ионы распределены неравномерно по обе стороны клеточной мембраны, во-первых, вследствие неодинаковой проницаемости клеточной мембраны для различных ионов, во-вторых, в результате работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки концентрационному и электрическому градиентам. По поводу определения «проницаемость» и «проводимость» единого мнения пока не сложилось.
Читайте также: