Регуляция положения белков в органеллах клеток
Обновлено: 17.05.2024
Многие белки синтезируются в неактивном виде (предшественники) и после схождения с рибосом подвергаются постсинтетическим структурным модификациям. Эти конформационные и структурные изменения полипептидных цепей получили название посттрансляционных изменений. Они включают удаление части полипептидной цепи (частичный протеолиз), ковалентное присоединение одного или нескольких низкомолекулярных лигандов, связывание между собой субъединиц олигомерного белка, приобретение белком нативной конформации (фолдинг).
При частичном протеолизе, например, неактивные предшественники секретируемых ферментов – зимогены – образуют активный фермент после расщепления по определенным участкам молекулы. Наглядным примером последовательного протеолиза служит и образование активных форм инсулина или глюкагона из препрогормонов.
В ходе ковалентных модификаций структурные белки и ферменты могут активироваться или инактивироваться в результате присоединения различных химических групп: фосфатных, ацильных, метильных, олигосахаридных и др. Многочисленным модификациям подвергаются боковые радикалы некоторых аминокислот: в тиреоглобулине йодируются остатки тирозина, в факторах свертывания крови карбоксилируются остатки глутамата, в цепях тропоколлагена гидроксилируются остатки пролина и лизина.
У некоторых белков на N-конце имеются короткие последовательности гидрофобных аминокислотных остатков, которые называют сигнальными последовательностями. Эти участки играют важную роль в транспорте белков через мембраны. В процессе переноса через мембрану сигнальная последовательность отщепляется сигнальной пептидазой. В итоге белок приобретает функциональную активность, оказавшись в соответствующей органелле или вне клетки.
Существование посттрансляционной модификации расширяет возможности клеток в регуляции метаболизма. Изменения количества или активности ферментов, участвующих в модификации белков, приводят к снижению или увеличению концентрации последних, что отражается на скорости соответствующих процессов.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Глава 2. Молекулярная архитектура белков
Глава 2. Молекулярная архитектура белков Не будем скрывать: покончив с первой главой, авторы (а возможно, и читатель) испытали некоторое облегчение. В конце концов цель ее заключалась лишь в том, чтобы дать читателю сведения, необходимые для понимания последующих глав,
Генеалогическое древо (свидетельство белков)
Генеалогическое древо (свидетельство белков) Генеалогическое древо (свидетельство белков)Различия в белках двух видов отражают эволюционные изменения этих видов после их отделения от общего предка. Анализ показывает, что между альбуминами кровяных сывороток шимпанзе
Вселенная древних белков продолжает расширяться
Вселенная древних белков продолжает расширяться В 2010 году журнал Nature опубликовал интересную статью об эволюционном движении белков по ландшафтам приспособленности (Povolotskaya, Kondrashov, 2010). Авторы работы решили сравнить аминокислотные последовательности 572 древних белков,
Изменения белков[43]
Изменения белков[43] Те участки генома, которые кодируют белки, изменились на удивление мало. Различия в аминокислотных последовательностях белков у человека и шимпанзе составляют значительно менее 1 %, да и из этих немногочисленных различий большая часть либо не имеет
Изменения активности генов
Изменения активности генов Эволюция животных в целом и приматов в частности протекает не столько за счет изменения структуры белок-кодирующих генов, сколько за счет изменения их активности. Небольшое изменение в верхних этажах иерархически организованных
Глава 2. Строение и функции белков
Глава 2. Строение и функции белков Белки – высокомолекулярные азотсодержащие органические соединения, состоящие из аминокислот, соединенных в полипептидные цепи с помощью пептидных связей, и имеющие сложную структурную организацию.История изучения белковВ 1728 г.
Уровни структурной организации белков
Уровни структурной организации белков Первичная структура – строго определенная линейная последовательность аминокислот в полипептидной цепочке.Стратегические принципы изучения первичной структуры белка претерпевали значительные изменения по мере развития и
Функционирование белков
Функционирование белков Каждый индивидуальный белок, имеющий уникальную первичную структуру и конформацию, обладает и уникальной функцией, отличающей его от всех остальных белков. Набор индивидуальных белков выполняет в клетке множество разнообразных и сложных
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Переваривание белков в желудочно-кишечном тракте
Переваривание белков в желудочно-кишечном тракте Переваривание белков начинается в желудке под действием ферментов желудочного сока. За сутки его выделяется до 2,5 литров и он отличается от других пищеварительных соков сильно кислой реакцией, благодаря присутствию
Расщепление белков в тканях
Расщепление белков в тканях Осуществляется с помощью протеолитических лизосомальных ферментов катепсинов. По строению активного центра выделяют цистеиновые, сериновые, карбоксильные и металлопротеиновые катепсины. Роль катепсинов:1. создание биологически активных
Роль печени в обмене аминокислот и белков
Роль печени в обмене аминокислот и белков Печень играет центральную роль в обмене белков и других азотсодержащих соединений. Она выполняет следующие функции:1. синтез специфических белков плазмы: - в печени синтезируется: 100 % альбуминов, 75 – 90 % ?-глобулинов, 50 %
Характеристика белков сыворотки крови
Характеристика белков сыворотки крови Белки системы комплемента – к этой системе относятся 20 белков, циркулирующих в крови в форме неактивных предшественников. Их активация происходит под действием специфических веществ, обладающих протеолитической активностью.
9.4.1. Циклические изменения
9.4.1. Циклические изменения Циклические изменения сообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов.Суточные преобразования в биоценозах обычно выражены тем сильнее, чем значительнее разница
8.2.1. Изменения климата
8.2.1. Изменения климата В настоящее время ни одну из рассмотренных версий происхождения Позднепермского и Позднетриасского кризисов (внутренние причины или удар астероида) нельзя считать доказанной окончательно. Однако осознание того, что и без вмешательства со стороны
Изменения в науке
Изменения в науке Суть научной деятельности не совсем такова, как она представляется нам. Мы привыкли считать, что развитие науки – это прямая дорога к постижению научных истин, путь, на котором постепенно накапливаются новые знания и расширяется круг рассматриваемых
Регуляция положения белков в органеллах клеток
Транспорт белков через эндоплазматический ретикулум (ЭПР) и аппарат Гольджи
Основные положения:
• Все белки, локализованные в ЭПР, аппарате Гольджи или в плазматической мембране, при синтезе вначале связываются с ЭПР.
• Белки переносятся из одного компартмента в другой при участии мембранных везикул, которые отпочковываются от поверхности одной мембраны и сливаются с поверхностью следующей.
• Транспорт белков из окружающей среды в клетку происходит в обратном направлении при участии везикул.
Интеграция белка в мембрану или его трансмембранный транспорт являются своеобразными событиями. Если белок должен пройти через несколько мембран, то его связывание с мембраной происходит на начальном этапе процесса транспорта. После этого он остается в мембранном окружении и транспортируется от одной мембраны к другой в везикуле.
Это, например, характерно для белков, исходно ассоциированных с ЭПР, но в конце высвобождающихся из плазматической мембраны. Аналогичная система используется для транспортировки белков как из клетки, так и в клетку.
Экзоцитоз представляет собой процесс, посредством которого белки транспортируются к плазматической мембране или во внеклеточную среду (обзор путей экзоцитоза).
Некоторые белки секретируются конститутивно, т. е. после синтеза они всегда выходят из клетки. Другие, продуцируемые некоторыми специализированными клетками, например пищеварительные ферменты, выходят только после получения клеткой соответствующего сигнала.
При попадании в ЭПР белок остается в мембране или в люмене до тех пор, пока он не переместится в другую часть системы с участием везикул, или не поступит обратно в цитозоль, где подвергнется деградации.

Перенос белков между компартментами,
ограниченными мембраной, происходит при отпочковывании от одного компартмента везикулы,
содержащей белки, и последующим ее слиянием с мембраной другого компартмента.
Транспортные везикулы представляют собой мелкие пузырьки (обычно 100-200 нм в диаметре), которые образуются при «отпочковывании» от мембраны (представления о везикулярном транспорте белков). Пузырек отпочковывается от поверхности одной мембраны и мигрирует к другой мембране, с которой он сливается.
Транспортные пузырьки называются окаймленными везикулами, поскольку их мембрана окружена белковой оболочкой. Различные типы везикул различаются между собой по составу оболочки, которая участвует в позиционировании и отборе транспортируемых белков.
Растворимые белки переносятся из донорного в акцепторный компартмент, находясь внутри везикулы; а мембранные белки переносятся в составе мембраны везикулы. Это означает, что белок может транспортироваться из ЭПР в аппарат Гольджи сквозь стопки Гольджи и к плазматической мембране.
Импорт белков в клетку также происходит с участием окаймленных везикул. Они образуются на плазматической мембране и поступают в клетку; этот процесс носит название эндоцитоз. Эндоцитоз происходит по тому же механизму, что и экзоцитоз, но протекает в обратном направлении.

Транспортные везикулы перемещают белки через ЭПР,
аппарат Гольджи и плазматическую мембрану.
Везикулы, которые отпочковываются от плазматической мембраны, включают материал из внеклеточной среды и возвращают его в плазматическую мембрану. Везикулы, которые осуществляют эндоцитоз, отличаются от везикул экзоцитоза составом белковой оболочки.
Некоторые патогенные микроорганизмы поникают в клетку хозяина посредством эндоцитоза; действительно, митохондрии и хлоропласта своим происхождением обязаны эндоцитозу. На рисунке ниже представлены аналогичные механизмы для транспорта вновь синтезированных белков по механизму экзоцитоза и для импорта белков в клетку по механизму эндоцитоза.
В каждом случае белок может транспортироваться при прохождении серии событий отпочковывания и слияния по мере того, как он мигрирует с поверхности одной мембраны на другую.
Один из результатов этого процесса состоит в том, что по мере протекания процессов отпочковывания и слияния происходит постоянное перемещение компонентов от одной мембраны к другой. С количественной точки зрения, при экзоцитозе переносится больше материала, чем при эндоцитозе, и прямой (антероградный) транспорт приводит к сплошному потоку липидов от ЭПР к плазматической мембране. За счет ретроградного транспорта, пустые везикулы возвращают липиды обратно в мембраны. Это обеспечивает целостность мембран.
Везикулярный транспорт белков представляет собой высокоспецифичный процесс. Основной принцип отбора состоит в том, что белок узнается по наличию специфического сигнала (чаще всего это короткая последовательность аминокислот) и включается в везикулу для дальнейшего транспорта. При отсутствии такого сигнала белок остается в компартменте, в котором он находится, или же продвигается медленнее. Отпочковывающиеся везикулы могут селективно включать белки, подлежащие транспорту, оставляя на местах резидентные белки органеллы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Энергетический обмен
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза - диссимиляции и ассимиляции, постоянно протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться) количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция - от лат. dissimilis ‒ несходный) - обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров, белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Подготовительный этап осуществляется ферментами в ЖКТ. В результате действия ферментов сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры - на глицерин и жирные кислоты, сложные углеводы - до простых сахаров.

Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК). Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов - организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на этапе гликолиза, синтезируется 18 молекул АТФ - в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.

АТФ - аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ - универсального источника энергии. Молекула АТФ состоит из азотистого основания - аденина, углевода - рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи - ковалентные связи, которые гидролизуются с выделением большого количества энергии. Их принято обозначать типографическим знаком тильда "∽".

- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции), удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Органоиды клетки
Органоиды (органеллы) клетки - специализированные структуры клетки, выполняющие различные жизненно необходимые функции. Особенно сложно устроены клетки простейших, где одна клетка составляет весь организм и выполняет функции дыхания, выделения, пищеварения и многие другие.
- Немембранные - рибосомы, клеточный центр, микротрубочки, органоиды движения (жгутики, реснички)
- Одномембранные - ЭПС, комплекс (аппарат) Гольджи, лизосомы и вакуоли
- Двумембранные - ядро, пластиды, митохондрии

Прежде чем говорить об органоидах клетки, без которых невозможна ее жизнедеятельность, необходимо упомянуть о том, без чего вообще не существует клетки - о клеточной мембране. Клеточная мембрана ограничивает клетку от окружающего мира и формирует ее внутреннюю среду.
Клеточная мембрана (оболочка)
Запомните, что в отличие от клеточной стенки, которая есть только у растительных клеток и у клеток грибов (она придает им плотную, жесткую форму) клеточная мембрана есть у всех клеток без исключения! Этот важный момент объясню еще раз :) У клеток животных имеется только клеточная мембрана, а у клеток растений и грибов есть и клеточная стенка, и клеточная мембрана.
Клеточная мембрана представляет собой билипидный слой (лат. bi - двойной + греч. lipos - жир), который пронизывают молекулы белков.

Билипидный слой представлен двумя слоями фосфолипидов. Обратите внимание, что их гидрофобные концы обращены внутрь мембраны, а гидрофильные "головки" смотрят наружу. Билипидный слой насквозь пронизывают интегральные белки, частично - погруженные белки, имеются также поверхностно лежащие белки - периферические.
- Поддержании постоянства структуры мембраны
- Рецепции сигналов из окружающей среды (химического раздражения)
- Транспорте веществ через мембрану
- Ускорении (катализе) реакций, которые ассоциированы с мембраной
Интегральные (пронизывающие) белки образуют каналы, по которым молекулы различных веществ могут поступать в клетку или удаляться из нее. "Заякоренные" молекулы олигосахаридов на поверхности клетки образуют гликокаликс, который выполняет рецепторную функцию, участвует в избирательном транспорте веществ через мембрану.

Теперь вы знаете, что гликокаликс - надмембранный комплекс, совокупность клеточных рецепторов, которые нужны клетке для восприятия регуляторных сигналов биологически активных веществ (гормонов, гормоноподобных веществ). Гормон избирателен, специфичен и присоединяется только к своему рецептору: меняется конформация молекулы рецептора и обмен веществ в клетке. Так гормоны регулируют жизнедеятельность клеток.
Вирусы и бактерии не являются исключением: они взаимодействуют только с теми клетками, на которых есть подходящие к ним рецепторы. Так, вирус гриппа поражает преимущественно клетки слизистой верхних дыхательных путей. Однако, если рецепторов нет, то вирус не может проникнуть в клетку, и организм приобретает невосприимчивость к инфекции. Вспомните врожденный иммунитет: именно по причине отсутствия рецепторов человек не восприимчив ко многим болезням животных.

Итак, вернемся к клеточной мембране. Ее можно сравнить со стенами помещения, в котором, вероятно, вы находитесь. Стены дома защищают его от ветра, дождя, снега и прочих факторов внешней среды. Рискну предположить, что в вашем доме есть окна и двери, которые по мере необходимости открываются и закрываются :) Так и клеточная мембрана может сообщать внутреннюю среду клетки с внешней средой: через мембрану вещества поступают в клетку и удаляются из нее.
- Разделительная (барьерная) - образует барьер между внешней средой и внутренней средой клетки (цитоплазмой с органоидами)
- Поддержание обмена веществ между внешней средой и цитоплазмой
Через мембрану по каналам кислород и питательные вещества поступают в клетку, а продукты жизнедеятельности - мочевина - удаляются из клетки во внешнюю среду.
- Пассивный - часто идет по градиенту концентрации, без затрат АТФ (энергии). Возможен путем осмоса, простой диффузии или облегченной (с участием белка-переносчика) диффузии.
Внутрь клетки с помощью осмоса поступает вода. Путем простой диффузии в клетку попадают O2, H2O, CO2, мочевина. Облегченная диффузия характерна для транспорта глюкозы, аминокислот.
Активный транспорт чаще происходит против градиента концентрации, в ходе него используются белки-переносчики и энергия АТФ. Ярким примером является натрий-калиевый насос, который накачивает ионы калия внутрь клетки, а ионы натрия выводит наружу. Это происходит против градиента концентрации, поэтому без затрат энергии (АТФ) не обойтись.

- Фагоцитоз (греч. phago - ем + cytos - клетка) - поглощение твердых пищевых частиц и бактерий фагоцитами
- Пиноцитоз (греч. pino - пью) - поглощение клеткой жидкости, захват жидкости клеточной поверхностью
Фагоцитоз был открыт И.И. Мечниковым, который создал фагоцитарную теорию иммунитета. Это теория гласит, что в основе иммунной системы нашего организма лежит явление фагоцитоза: попавшие в организм бактерии уничтожаются фагоцитами (T-лимфоцитами), которые переваривают их.
В ходе эндоцитоза мембрана сильно прогибается внутрь клетки, ее края смыкаются, захватывая бактерию, пищевые частицы или жидкость внутрь клетки. Образуется везикула (пузырек), который движется к пищеварительной вакуоли или лизосоме, где происходит внутриклеточное пищеварение.

Клетки многих органов, к частности эндокринных желез, которые выделяют в кровь гормоны, транспортируют синтезированные вещества к мембране и удаляют их из клетки с помощью экзоцитоза (от др.-греч. ἔξω - вне, снаружи). Таким образом, процессы экзоцитоза и эндоцитоза противоположны.
Клеточная стенка
Расположена снаружи клеточной мембраны. Присутствует только в клетках бактерий, растений и грибов, у животных отсутствует. Придает клетке определенную форму, направляет ее рост, придавая характерное строение всему организму. Клеточная стенка бактерий состоит из полимера муреина, у грибов - из хитина, у растений - из целлюлозы.

Цитоплазма
Органоиды клетки расположены в цитоплазме, которая состоит из воды, питательных веществ и продуктов обмена. В цитоплазме происходит постоянный ток веществ: поступившие в клетку вещества для расщепления необходимо доставить к органоидам, а побочные продукты - удалить из клетки.
Постоянное движение цитоплазмы поддерживает связь между органоидами клетки и обеспечивает ее целостность.

Прокариоты и эукариоты
Прокариоты (греч. πρό - перед и κάρυον - ядро) или доядерные - одноклеточные организмы, не обладающие в отличие от эукариот оформленным ядром и мембранными органоидами. У прокариот могут обнаруживаться только немембранные органоиды. Их генетический материал представлен в виде кольцевой молекулы ДНК - нуклеоида (нуклеоид - ДНК–содержащая зона клетки прокариот). К прокариотам относятся бактерии, в их числе цианобактерии (цианобактерий по-другому называют - сине-зеленые водоросли).
Эукариоты (греч. εὖ - хорошо + κάρυον - ядро) или ядерные - домен живых организмов, клетки которых содержат оформленное ядро. Растения, животные, грибы - относятся к эукариотам.

Немембранные органоиды
Очень мелкая органелла (около 20 нм), которая была открыта после появления электронного микроскопа. Состоит из двух субъединиц: большой и малой, в состав которых входят белки и рРНК (рибосомальная РНК), синтезируемая в ядрышке.
Запомните ассоциацию: "Рибосома - фабрика белка". Именно здесь в ходе матричного биосинтеза - трансляции, с которой подробнее мы познакомимся в следующих статьях, на базе иРНК (информационной РНК) синтезируется белок - последовательность соединенных аминокислот в заданном иРНК порядке.

Микротрубочки являются внутриклеточными белковыми производными, входящими в состав цитоскелета. Они поддерживают определенную форму клетки, участвуют во внутриклеточном транспорте и процессе деления путем образования нитей веретена деления. Микротрубочки также образуют основу органоидов движения: жгутиков (у бактерий жгутик состоит из сократительного белка - флагеллина) и ресничек.
Микрофиламенты - тонкие длинные нитевидные структуры, состоящие из белка актина. Встречаются во всей цитоплазме, служат для создания тока цитоплазмы, принимают участие в движении клетки, в процессах эндо- и экзоцитоза.

Этот органоид характерен только для животной клетки, в клетках грибов и высших растений отсутствует. Клеточный центр состоит из 9 триплетов микротрубочек (триплет - три соединенных вместе). Участвует в образовании нитей веретена деления, располагается на полюсах клетки.

Это органоиды движения, которые выступают над поверхностью клетки и имеют в основе пучок микротрубочек. Реснички встречаются только в клетках животных, жгутики можно обнаружить у животных, растений и бактерий.
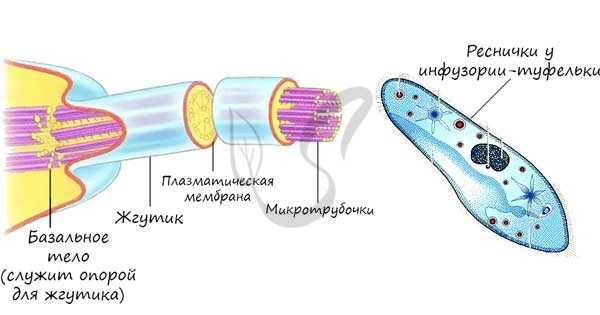
Одномембранные органоиды
- Эндоплазматическая сеть (ЭПС), эндоплазматический ретикулум (лат. reticulum - сеть)
ЭПС представляет собой систему мембран, пронизывающих всю клетку и разделяющих ее на отдельные изолированные части (компартменты). Это крайне важно, так как в разных частях клетки идут реакции, которые могут помешать друг другу, что нарушит процессы жизнедеятельности.
Выделяют гладкую ЭПС и шероховатую ЭПС. Обе они выполняют функцию внутриклеточного транспорта веществ, однако между ними имеются различия. На мембранах гладкой ЭПС происходит синтез липидов, обезвреживаются вредные вещества. Шероховатая ЭПС синтезирует белок, так как имеет на мембранах многочисленные рибосомы (потому и называется шероховатой).

Комплекс Гольджи состоит из трубочек, сети уплощенных канальцев (цистерн) и связанных с ними пузырьков. Располагается вокруг ядра клетки, внешне напоминает стопку блинов. Это - "клеточный склад". В нем запасаются жиры и углеводы, с которыми здесь происходят химические видоизменения.
Модифицированные вещества упаковываются в пузырьки и могут перемещаться к мембране клетки, соединяясь с ней, они изливают свое содержимое во внешнюю среду. Можно догадаться, что комплекс Гольджи хорошо развит в клетках эндокринных желез, которые в большом количестве синтезируют и выделяют в кровь гормоны.
В комплексе Гольджи появляются первичные лизосомы, которые содержат ферменты в неактивном состоянии.

Представляет собой мембранный пузырек, содержащий внутри ферменты (энзимы) - липазы, протеазы, фосфатазы. Лизосому можно ассоциировать с "клеточным желудком".
Лизосома участвует во внутриклеточном пищеварении поступивших в клетку веществ. Сливаясь с фагосомой, первичная лизосома превращается во вторичную, ферменты активируются. После расщепления веществ образуется остаточное тельце - вторичная лизосома с непереваренными остатками, которые удаляются из клетки.

Лизосома может переварить содержимое фагосомы (самое безобидное), переварить часть клетки или всю клетку целиком. В норме у каждой клетки жизненный цикл заканчивается апоптозом - запрограммированным процессом клеточной гибели.
В ходе апоптоза ферменты лизосомы изливаются внутрь клетки, ее содержимое переваривается. Предполагают, что нарушение апоптоза в раковых клетках ведет к бесконтрольному росту опухоли.

Пероксисомы (микротельца) содержат окислительно-восстановительные ферменты, которые разлагают H2O2 (пероксид водорода) на воду и кислород. Если бы пероксид водорода оставался неразрушенными, это приводило бы к серьезным повреждениям клетки.
Вакуоли характерны для растительных клеток, однако встречаются и у животных (у одноклеточных - сократительные вакуоли). У растений вакуоли выполняют другие функции и имеют иное строение: они заполняются клеточным соком, в котором содержится запас питательных веществ. Снаружи вакуоль окружена тонопластом.
Трудно переоценить значение вакуолей в жизнедеятельности растительной клетки. Вакуоли создают осмотическое давление, придают клетке форму.
Примечательно, что по размеру вакуолей можно судить о возрасте клетки: молодые клетки имеют вакуоли небольшого размера, а в старых клетках вакуоли могут настолько увеличиваться, что оттесняют ядро и остальные органоиды на периферию.

Двумембранные органоиды
Важнейший компонент эукариотической клетки - оформленное ядро, которое у прокариот отсутствует. Внутренняя часть ядра представлена кариоплазмой, в которой расположен хроматин - комплекс ДНК, РНК и белков, и одно или несколько ядрышек.
Ядрышко - место в ядре, где активно идет процесс матричного биосинтеза - транскрипция, с которым мы познакомимся подробнее в следующих статьях. В течение дня, наблюдая за одной и той же клеткой, можно увидеть разное количество ядрышек или не найти ни одного.

Замечу, что хромосомы видны только в момент деления клетки. Хромосомы представляют собой сильно спирализованные молекулы ДНК, связанные с белками.
Я всегда рекомендую ученикам ассоциировать хромосому с мотком ниток: если все нитки обмотать вокруг одной оси, то они становятся мотком и хорошо видны (хромосомы - во время деления, спирализованное ДНК), если же клетка не делится, то нитки размотаны и разбросаны в один слой, хромосом не видно (хроматин - деспирализованное ДНК).

Хромосомы отличаются друг от друга по строению, форме, размерам. Совокупность всех признаков (форма, число, размер) хромосом называется кариотип. Кариотип может быть представлен по-разному: существует кариотип вида, особи, клетки.
Изучая кариотип человека, врач-генетик может обнаружить различные наследственные заболевания, к примеру, синдром Дауна - трисомия по 21-ой паре хромосом (должно быть 2 хромосомы, однако при синдроме Дауна их три).

Органоид палочковидной формы. Митохондрию можно сравнить с "энергетической станцией". Если в цитоплазме происходит анаэробный этап дыхания (бескислородный), то в митохондрии идет более совершенный - аэробный этап (кислородный). В результате кислородного этапа (цикла Кребса) из двух молекул пировиноградной кислоты (образовавшихся из 1 глюкозы) получаются 36 молекул АТФ.
Митохондрия окружена двумя мембранами. Внутренняя ее мембрана образует выпячивания внутрь - кристы, на которых имеется большое скопление окислительных ферментов, участвующих в кислородном этапе дыхания. Внутри митохондрия заполнена матриксом.

Запомните, что особенностью этого органоида является наличие кольцевой молекулы ДНК - нуклеоида (ДНК–содержащая зона клетки прокариот), и рибосом. То есть митохондрия обладает собственным генетическим материалом и возможностью синтеза белка, почти как отдельный организм.
В связи с этим, митохондрия считается полуавтономным органоидом. Вероятнее всего, изначально митохондрии были самостоятельными организмами, однако со временем вступили в симбиоз с эукариотами и стали частью клетки.
Митохондрий особенно много в клетках мышц, в том числе - в сердечной мышечной ткани. Эти клетки выполняют активную работу и нуждаются в большом количестве энергии.
- Хлоропласт (греч. chlōros - зелёный)
Получил свое название за счет содержащегося в нем зеленого пигмента - хлорофилла (греч. chloros - зеленый и phyllon - лист). Под двойной мембраной расположены тилакоиды, которые собраны в стопки - граны. Внутреннее пространство между тилакоидами и мембраной называется стромой.
Запомните, что светозависимая (световая) фаза фотосинтеза происходит на мембранах тилакоидов, а темновая (светонезависимая) фаза - в строме хлоропласта за счет цикла Кальвина. Это очень пригодится при изучении фотосинтеза в дальнейшем.

Так же, как и митохондрии, пластиды относятся к полуавтономным органоидам: в них имеется кольцевидная ДНК (находится в нуклеоиде), рибосомы.
Пластиды, которые содержат пигменты каратиноиды в различных сочетаниях. Сочетание пигментов обуславливает красную, оранжевую или желтую окраску. Находятся в плодах, листьях, лепестках цветков.
Хромопласты могут развиваться из хлоропластов: во время созревания плодов хлоропласты теряют хлорофилл и крахмал, в них активируется биосинтез каротиноидов.
Не содержат пигментов, образуются в запасающих частях растения (клубни, корневища). В лейкопластах накапливается крахмал, липиды (жиры), пептиды (белки). На свету лейкопласты могут превращаться в хлоропласты и запускать процесс фотосинтеза.

Посттрансляционные изменения белков
Многие белки синтезируются в неактивном виде (предшественники) и после схождения с рибосом подвергаются постсинтетическим структурным модификациям. Эти конформационные и структурные изменения полипептидных цепей получили название посттрансляционных изменений. Они включают удаление части полипептидной цепи (частичный протеолиз), ковалентное присоединение одного или нескольких низкомолекулярных лигандов, связывание между собой субъединиц олигомерного белка, приобретение белком нативной конформации (фолдинг).
Регуляция синтеза белка
Соматические клетки всех тканей и органов многоклеточного организма содержат одинаковую генетическую информацию, но отличаются друг от друга по содержанию тех или иных белков. Для эритроцитов, например, характерно высокое содержание гемоглобина, для клеток соединительной ткани – коллагена, клетки поджелудочной железы вырабатывают много ферментов. В отдельных клетках, тканях и органах содержание разных белков меняется онтогенез. Все это свидетельствует о том, что в живых организмах существуют механизмы, регулирующие белковый синтез. Они функционируют под действием внутренних и внешних факторов на каждой из стадий сложного процесса синтеза белка. Количество протеинов может изменяться в результате увеличения числа некоторых генов, регуляции на стадии транскрипции, процессинга мРНК. Скорость белкового синтеза определяется также и временем жизни мРНК, регуляцией синтеза на уровне трансляции и посттрансляционной модификации белков.
Регуляция на самых ранних этапах (на уровне экспрессии генов) является наиболее выгодной и поэтому широко встречается у эукариотических организмов. На экспрессию генов у эукариот влияет целый ряд факторов.
Организация хроматина и доступность генов: в ядрах дифференцированных клеток хроматин имеет такую укладку, что только небольшое число генов доступно для транскрипции. Различают участки гетерохроматина, в которых ДНК упакована очень компактно и для транскрипции недоступна, и участки эухроматина, имеющие более рыхлую укладку и способные связывать РНК-полимеразу. В разных типах клеток в область эухроматина попадают разные гены. Это ведет к тому, что в разных тканях транскрибируются разные участки хроматина.
Изменение количества генов: амплификация (увеличение числа) генов при необходимости увеличения синтеза определенного генного продукта; утрата генетического материала (процесс, происходящий при созревании некоторых типов клеток, например, эритроцитов).
Перестройка генов или генетичесая рекомбинация: перемещение генов между хромосомами или внутри одной хромосомы, объединение генов с образованием измененной хромосомы, которая после таких изменений способна к репликации и транскрипции.
Регуляция транскрипции (см. лекцию № 6).
Существенное значение в обеспечении разнообразия белков играет посттранскрипционный процессинг РНК. Основные способы такой регуляции – альтернативный сплайсинг и изменение стабильности РНК.
Известны и некоторые случаи регуляции количества и разнообразия белков путем изменения скорости процесса их трансляции. Наиболее изученный пример – синтез белков в ретикулоцитах. Известно, что на этом уровне дифференцировки кроветворные клетки лишены ядра, а следовательно и ДНК. Регуляция синтеза белка-глобина осуществляется только на уровне трансляции и зависит от содержания гема в клетке.
Читайте также:
- Иммуногистология заболевания пародонта. Функциональная активность ПМЯЛ
- Пневмоцистные инфекции - причины, патогенез
- Симптомы лимфангиомы шеи (кистозной гигромы) и ее лечение
- Почечный кровоток и потребление кислорода. Факторы влияющие на почечный кровоток
- Диагностика цирроза печени. Лечение портальной гипертензии.